Share on Facebook
Share on Facebook Share on X
Share on X Share by Email
Share by Email Share on Google Classroom
Share on Google Classroom

Structure
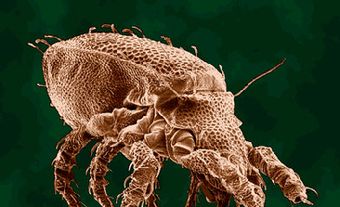
Bees are characterized by branched hairs on the body, chewing mouthparts accompanied by a long proboscis (or tongue) for drinking nectar, transparent wings with few veins, and hind legs that are often enlarged with stiff hairs for gathering pollen. Almost all species are adapted for gathering nectar and pollen from blossoms, although males do not actively gather pollen, and therefore usually lack the specialized pollen-carrying structures of females.
Eusocial colonies are made up of three different castes: queens (fertile and egg-laying females), workers (sterile female offspring of the queen) and drones (males). Queens are usually larger than workers and drones and have enlarged abdomens, while workers are slightly smaller than drones. Bees range from 2 mm to 4 cm in length1, and greatly vary in colour, with species that are black, brown, yellow, white, orange, red, grey, metallic shades of green, blue and purple, as well as many combinations of these colours.
Distribution and Habitat
Bees are most diverse in dry and Mediterranean climates, and especially so in warm deserts and shrublands. Bees prefer sunny habitats with an abundance of flowers, but can be found wherever the climate permits flowering plants to persist. Sub-Arctic and Arctic regions, however, support far fewer bees, and primarily small populations of larger-bodied bumble bees. There, much of the pollination needs of flowers are instead fulfilled by flies.
Pollination

The pollen of most flowering plants is collected and transported by bees, and most fruits, vegetables, and other important crops like flax, alfalfa and canola are bee-pollinated. Though bees are not the only pollinators of flowering plants — they share this role with birds, bats, moths and flies — they are undoubtedly the most important. Just how important they are to global biodiversity is impossible to estimate; non-bee-pollinated plants inevitably depend in part on bee-pollinated ones, such that a large proportion of any given ecosystem also depends indirectly on bees.
Different bee species exhibit different levels of specialization in terms of the flowers they visit. Honey bees are extreme generalists, and readily collect pollen from a huge variety of unrelated kinds of flowers (polylectic). Many bees are more specialized, however, and only collect pollen from a few closely related plant species (oligolectic). Many oligolectic bees have evolved morphological and behavioural adaptations that make them better at collecting pollen from — and therefore better at pollinating — their associated plants. Buzz-pollination is one such behaviour, whereby a flower is vibrated, or “buzzed,” to release otherwise inaccessible pollen. Many plants require this vibration to release their pollen (e.g., tomatoes), and a correspondingly broad range of bees exhibit this behaviour, although honey bees do not. Such differences in pollination efficiency have the result that, for many crops — including alfalfa, watermelon, blueberries and other berries — wild bees make much better pollinators than honey bees. In many cases, wild-bee-pollinated fruit crops can be economically viable without the use of managed honey bees, provided they are planted in close proximity to natural bee habitat, or if such habitat is actively incorporated into the crops.
Bees also collect nectar from flowers, which serves as food both for adults and, when mixed with pollen, for larvae. Some bees also provision their nests with floral oils (e.g., Macropis nuda), usually used in place of nectar for larval food.
Reproduction and Development
Sex is regulated by fertilization of the eggs: eggs that are fertilized by sperm as they are laid become female, while unfertilized eggs develop into males. This type of sex determination is called haplodiploidy because males remain haploid (with only half the number of chromosomes of diploid females).
In social bees, external secretions (pheromones), especially from the queen, regulate caste structure and behaviour, with the result that only queens lay eggs. Other pheromones are used to mark trails to food sources, although honey bees also communicate direction and distance to flowers or other resources using their “waggle dance.”
Nesting Biology
The nesting behaviour of bees ranges from completely solitary, to a few (often related) individuals sharing nests, to highly eusocial. Most bees are solitary. In fact, only an estimated 10 per cent of bee species are eusocial — meaning they establish nests that exhibit overlapping generations, co-operative brood care, and a caste system consisting of a queen, workers and drones. These colonies consist entirely of the founding queen and her offspring.
Solitary Bees

Most solitary bees excavate tunnels in soil where they construct nest cells, though some nest in holes they chew into wood (such as the carpenter bees, genus Xylocopa), or in the hollow centres of dead plant stems (e.g., genus Ceratina and genus Hylaeus). Many ground-nesting bees fashion unique clay nest cells underground (such as digger bees, genus Anthophora). Individual nest cells may be lined or not. Leafcutter bees (family Megachilidae), line their nest cells with cut pieces of leaves, carried back to the nest, while other megachilid bees line their nests with plant hairs or resin, or sometimes even pieces of synthetic materials like plastic shopping bags. Cellophane bees (family Colletidae) are so-named for the waterproof, cellophane-like material they secrete to line their nest cells. Females of all solitary species make their own nests in which they provision each cell with pollen and nectar, deposit their eggs, and leave offspring to develop independently.
Intermediate Levels of Sociality
Some bees are considered subsocial, meaning that females remain with the nest after their offspring have hatched, feeding and caring for them, as in some species of the small carpenter bee genus Ceratina. Others are communal, meaning multiple females use the same nest, but each provisions its own cells and lays its own eggs — this is the case for a number of species of sweat bees (family Halictidae). The degree of sociality may also be flexible within a species, as in the orange-legged furrow bee, Halictus rubicundus, which may be solitary or eusocial in different parts of its range or at different times of the year.
Eusocial Bees

In Canada, eusocial bees are represented by bumble bees, honey bees, and some sweat bees in the genera Halictus and Lasioglossum. Bumble bees are eusocial bees with annual colonies. Their nests comprise small, pot-like containers made from wax, used to store honey and pollen and to raise a brood. The queen emerges in spring, finds a nesting site (often in a rodent nest) to start a colony and is actively seen foraging for floral provisions. She provisions the first batch of eggs she produces in the spring, which develop into workers. As summer progresses workers take over the roles of foraging and feeding the brood. Later, the queen lays eggs that are destined to become new queens and drones. Colonies usually contain fewer than a few hundred to a thousand individuals, depending on the species. The social structure of the colony breaks down as the new queens are mated and each one seeks an underground shelter to spend the winter. Only these new and mated queens survive until the next year. In Canada, 41 free-nesting bumble bee species are found, several above the Arctic Circle.
The most widely studied eusocial bee is the honey bee. The precise architecture of the hexagonal cells of the comb, the complex social behaviour of the colony, the honey and beeswax they produce, and the pollination they provide make honey bees among the most fascinating and useful insects to humans. The colonies are mostly highly populous, with tens of thousands of individual bees under the influence of a single queen. A colony’s population fluctuates during the year, being highest in late spring and early summer when swarming is most likely to occur, and lowest in late winter. The colony is active all winter long, kept warm by the bees’ own body heat. The brood requires a temperature of about 35°C to develop properly. Swarming is the colony’s way of reproducing. A mass of thousands of bees, with a queen, exits from the parent colony and finds a new home. The parent colony continues in the old home with a new queen.
Parasitic Bees
Some bees have evolved into parasites of their relatives and have lost the ability to gather food and rear their own offspring. Some are social parasites, such as bumble bees of the subgenus Psithyrus, whose queens seek out established bumble bee nests and either replace or live alongside the original queen. The parasitic queen lays eggs that the host workers rear, producing only reproductive queens and drones. Most parasitic bees are cleptoparasites, commonly called cuckoo bees because, like cuckoo birds, they lay their eggs in the already-established nests of their hosts. The parasitic larvae hatch and eat the food intended for host larvae, while the latter are killed by the cuckoos.
Bee Stings
Only female bees have a sting, which is a repurposed egg-laying organ, or ovipositor, used exclusively for defence against would-be predators (bees, along with stinging wasps and ants, instead lay eggs through a separate opening). The sting is found at the tip of the female’s abdomen and is used to inject a toxic venom that is responsible for the pain, swelling and sometimes more severe allergic reactions that result from a sting. Only about 1-3 per cent of the general population exhibit life-threatening (i.e., anaphylactic) allergic responses to bee and wasp stings.
In the case of honey bees, the workers’ sting is barbed. When the bee stings a thick-skinned victim such as a human, a part of its abdomen is usually torn off and left behind, causing death. The stings of most other bees are not so-barbed, allowing them to use their stings again and again. This includes honey bee queens, which, along with queens of other eusocial colonies, use their stings only against other queens. While honey bees may sting people that approach their hives too closely, most bees are not aggressive, and normally only sting humans if trapped or pressed against the skin.
The so-called “stingless bees” (tribe Meliponini of the family Apidae) of the tropics and subtropics have only rudimentary stings that are useless for defence. These bees resort to other means of colony protection, such as biting and applying irritant plant materials to their foes. Many small, commonly encountered bees (e.g., Lasioglossum sweat bees) have very small stings that often fail to break skin; when they do, the sting is usually no more painful than a pinprick.
Environmental Threats and Concerns
Fifteen of Canada’s wild bee species (all but two of which are bumble bees) are listed as threatened by various bodies. The International Union for Conservation of Nature lists three species as critically endangered, and six as vulnerable13. Only one species, the rusty-patched bumble bee (Bombus affinis), is protected under the federal Species at Risk Act as endangered. Others are listed under various provincial and territorial species-at-risk frameworks. Among these, the most commonly listed species are the gypsy cuckoo bumble bee (Bombus bohemicus, endangered) and the yellow-banded bumble bee (Bombus terricola, special concern). Along with the rusty-patched bumble bee, these bees were once common across much of North America, but have declined rapidly since the 1990s. The last confirmed sighting in Canada of the rusty-patched bumble bee was in 2009, in Pinery Provincial Park, Ontario.
There are a number of identified threats to the health of bee populations, in particular for wild bee species. These threats include: habitat loss, insecticides, competition with introduced bees and pathogen spillover from managed bees. Habitat loss is mainly the result of intensified agriculture and urban development, leading to the loss of foraging and nesting habitat. Insecticides can negatively impact the survival of a population in the long term. Notably, neonicotinoid pesticides have recently been implicated in declines of wild bees, managed bees and honey bees alike. These types of pesticides not only reduce the ability of bee populations to survive through the winter, but also impair bees’ ability to learn, communicate, establish new nests, and forage effectively. In the case of competition, introduced bees, such as managed honey bees, can deplete resources that would otherwise be abundant for wild bees. Similarly, pathogens found among managed bees are a particular concern for wild bumble bee populations, which have been found to have a higher incidence of disease when foraging near greenhouses that use managed bumble bees.
Honey bees in particular have received a lot of attention due to the widespread phenomenon known as Colony Collapse Disorder (CCD). CCD is a complex problem, best understood as the outcome of a number of stressors that together cause the sudden loss of most of the adult population of a honey bee colony. These stressors include some of those discussed above, along with a number of parasites and diseases specific to honey bees. Colonies may be additionally stressed due to being shipped across the continent to wherever they’re needed to meet the pollination needs of large agricultural crops. In the process, transported honey bee colonies are exposed to a variety of pesticides, pathogens and pests, all the while being stressed nutritionally, as they are able to collect pollen and nectar from only a single plant species at a time.